6. La Ruta Pi3k/akt/mtor Y El Cáncer
6. La Ruta Pi3k/akt/mtor Y El Cáncer
La desregulación de la ruta mTOR es un elemento clave en el desarrollo de distintas enfermedades humanas y como consecuencia se están desarrollando drogas que tienen como diana dicha proteína (Figura 3). Una de las enfermedades más claramente asociadas con la desregulación de la ruta raptor-mTOR es el complejo de esclerosis tuberosa (TSC) y la linfangioleiomatosis (LAM) ambas causadas por mutaciones en los supresores de tumores TSC1 y TSC2. Pacientes con TSC desarrollan tumores benignos de crecimiento lento que cuando se presentan en el cerebro pueden causar ataques, retraso mental y muerte. LAM es una enfermedad rara, en la que los pacientes tienen seriamente comprometido el funcionamiento pulmonar, lo que resulta en una proliferación anormal de los fibroblastos pulmonares.
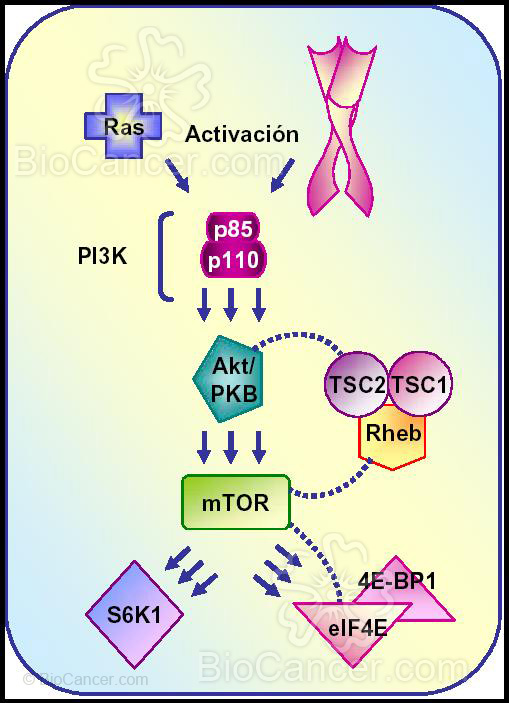
Figura 3: Desregulación de la ruta de señalización de PI·K/Akt/mTOR en humanos. La desregulación de esta ruta resulta de una activación endógena o exógena. Los factores exógenos incluyen la activación por Ras, principalmente restringido a tumores gastrointestinales., mientras que la avivación del receptor tirosinquinasa se ha descrito en una amplia variedad de tumores sólidos y hematológicos. Los factores endogenos incluyen bien la activación de quinasa por mutación génica/amplificación o por pérdida de la función de PTEN.
La activación constitutiva de esta ruta en las células cancerígenas tiene lugar en distintos niveles (Tabla 1). Los procesos intrínsecos que resultan en la activación de mTOR implican la pérdida de la función de PTEN, mutación o amplificación de la subunidad catalítica p110 de PI3K, la mutación de la subunidad reguladora de PI3K, p85, la amplificación de las isoenzimas de Akt, Akt1 y Akt2 y la inactivación o mutaciones de proteínas reguladoras de mTOR asociadas a Akt, como las proteínas TSC1 y TSC2. Además la ruta también puede ser activada vía oncogenes exógenos, que incluyen la elevada expresión o mutación de diferentes tipos de receptores tirosina quinasa, como el receptor del factor de crecimiento epidérmico (HER1-4), receptor del factor derivado de plaquetas (PDGFR)/KIT e IGFR, y Ras directamente unido a la subunidad p110 de PI3K. Por debajo de mTOR, la sobreexpresión y/o amplificación de S6K1 o eIF4E puede también contribuir a la oncogénesis. Sin embargo, no se han encontrado mutaciones en mTOR. Además, existe otra función de mTOR en la activación del cáncer. La activación de p53 es un regulador negativo de mTOR y, generalmente, la función de p53 se pierde de forma frecuente en cáncer, lo que provocaría una activación constitutiva de mTOR. El papel de estas proteínas transductoras de señales se ha estudiado profundamente en animales. Parece que la activación de esta ruta no es suficiente para inducir cáncer, necesitándose un evento oncogénico secundario para inducir la transformación.
La activación de la ruta PI3K/Akt/mTOR afecta a muchos tipos de tumores. Por ejemplo, la activación de PI3K mediada por mutaciones en K-Ras aparece en ciertos cánceres gastrointestinales y más particularmente en cánceres pancreáticos, gástricos y cáncer de colon. La pérdida de la función de PTEN bien por mutación, deleción o metilación del promotor, se ha descrito en cáncer de endometrio, glioblastoma (proceso neoplásico cerebral maligno), próstata, ovario, carcinoma de tiroides y en menos frecuencia en carcinoma hepático, mama, pulmón, carcinoma de las células renales y en melanoma.
|
Proteína |
Disfunción /Efecto |
Tipo de tumor |
|
K-Ras |
Mutación provoca activación |
Páncreas, colon, gástrico |
|
Receptor tirosina quinasa |
Activación de receptor |
Muchos tipos tumorales |
|
P110 |
Mutación génica Amplificación génica |
Cabeza y cuello, ovario Gastrointestinal, cerebro |
|
P85 |
Mutación génica |
Colon, ovario |
|
PTEN |
Mutación génica, deleción o metilación del promotor (perdida de función) |
Endometrial, glioblastoma, tiroides, carcinoma hepatocelular, síndrome de Cowden |
|
АКТ |
Amplificación génica Sobreexpresión proteica |
Mama, ovario, colon Ovario, mama |
|
TSC1/2 |
Mutación génica |
Síndrome TSC |
|
4EBP1 у eIF4E |
Amplificación génica Sobre-expresión proteica |
Mama Adenocarcinoma |
|
S6K1 |
Amplificación génica |
Mama, ovario |
Los tumores asociados a la pérdida de la función de PTEN son particularmente susceptibles a los inhibidores de mTOR. La inactivación de PTEN debida a alteraciones epigenéticas o genéticas puede resultar en la pérdida de la expresión de la proteína en varios tumores esporádicos:
La inhibición de la ruta raptor-mTOR en tumores sólidos dependerá del estado de activación de la ruta. Raptor-mTOR estará extremadamente activo en las zonas de tumores bien vascularizados bajo la estimulación de nutrientes y de factores de crecimiento derivados del tumor y del estroma. En estos casos, los inhibidores pueden ralentizar el crecimiento y la proliferación y quizás, de forma sinérgica con quimioterápicos, inducir la muerte celular. Por el contrario, en otras áreas pobremente irrigadas, la actividad raptor-mTOR será baja, debido a la ausencia de señales necesarias como nutrientes y oxígeno. En tales zonas, la supresión de raptor-mTOR disminuirá el crecimiento celular e inducirá a la autofagia, lo que permitirá a las células conservar la energía y nutrientes hasta que las condiciones ambientales mejoren. Por lo que un activador de raptor-mTOR podría ser terapéuticamente beneficioso, para conducir a las células a consumir la energía y los nutrientes, de manera que no puedan mantener los procesos vitales (48).