3. Bases Fisiológicas De La Acción De La Lh-rh
3. Bases Fisiológicas De La Acción De La Lh-rh
Una característica esencial de la secreción de todas hormonas hipotalámicas es su ritmo pulsátil. El caso de la regulación hipotalámica de la secreción de las dos gonadotrofinas, LH y FSH, mediante una sola hormona hipotalámica, LH-RH o Gn-RH, y por un tipo celular único, los gonadotrofos, es particularmente complejo. Las variaciones en la secreción relativa de FSH y LH durante la vida fértil se basan en la capacidad de los gonadotrofos hipofisarios de responder de forma variable a los pulsos secretores cambiantes de LH-RH al sistema portal hipotalámico hipofisario (1-4). La cantidad relativa de FSH y LH secretadas en cada momento del ciclo dependen de abundancia de receptores para LH-RH, y de la regulación de la respuesta intracelular a los pulsos de LH-RH de los gonadotrofos, influenciada por las hormonas ováricas estradiol y activina (Fig. 2).
LH-RH:
[pGlu1-His2-Trp3-Ser4-Tyr5-Gly6-Leu7-Arg8-Pro9-Gly10-NH2]
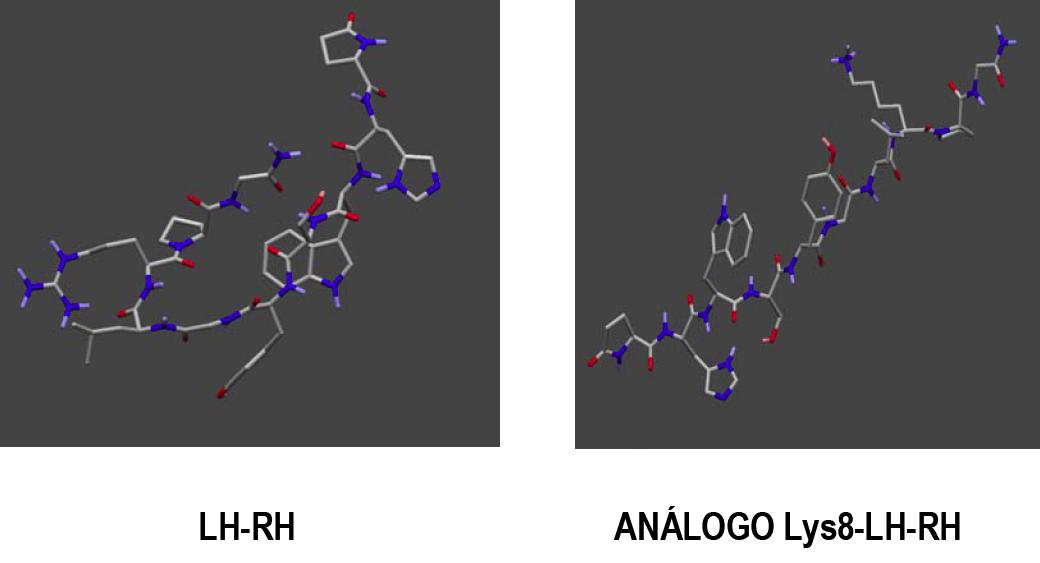
Figura 1.- ESTRUCTURA DE LH-RH Y UNO DE SUS ANÁLOGOS.
La figura muestra la secuencia de aminoácidos de la LH-RH y su estructura espacial. La
sustitución de Arg-8-Lys da lugar a una importante linearización de la estructura espacial,
que convierte al análogo en superagonista.
Una vez secretada la LH-RH al sistema portal hipotalámico-hipofisario, la hormona alcanza los gondotrofos hipofisarios. Debido al escaso flujo de sangre entre el hipotálamo y la hipófisis, la concentración de LH-RH es elevada en torno a esas células. Una vez que la sangre abandona la hipófisis para entrar en la circulación general, la LH-RH se diluye mucho, por lo que no parece capaz de estimular células extrahipofisarias. Por ello, a pesar de que existen muchas otras células en tejidos reproductivos que expresan receptores para LH-RH, seguramente los gonadotrofos constituyen la única diana fisiológica de la LH-RH, pues es en torno a ellos donde la concentración de hormona alcanza niveles de concentración suficientes para ocupar un número significativo de receptores.
El primer paso en la acción de la LH-RH comienza con su unión al receptor específico anclado en la membrana plasmática, lo que causa una microagregación de receptores y la formación de un complejo oligomérico (4). El complejo oligomérico hormona-receptor puede ahora ejercer su efecto señalizador.
El siguiente paso incluye la activación de una proteína G (intercambiadora de GTP), que a su vez activa la fosfolipasa C, que conduce a la producción de acilglicerol y a la activación de la proteínquinasa C. Como consecuencia de estos metabolitos se abren canales iónicos que incrementan la concentración de calcio intracelular, requisito necesario para la liberación de LH y FSH acumulada en gránulos de secreción intracelulares.

La respuesta de los gonadotrofos a la llegada pulsátil de LH-RH requiere la
internalización/destrucción del receptor en cada ciclo y su síntesis y recolocación
en la membrana, a la espera de una nueva oleada de hormona. El gen del receptor debe
pues responder a un conjunto de señales que condicionan su disponibilidad cíclica.
La figura muestra la compleja regulación de la expresión del gen, donde la autorregulación
juega el papel determinante, junto a la participación de la activina (ovárica e hipofisaria),
que probablemente modula su expresión para adaptarla a las necesidades secretoras
cambiantes a lo largo del ciclo de LH y FSH. La administración continuada de agonistas
desacopla el proceso y la síntesis de receptor no recibe las instrucciones adecuadas.
Finalmente los microagregados de receptores para LH-RH son internalizados y degradados por proteasas lisosomales intracelulares. La eliminación de receptores de LH-RH da lugar a la insensibilidad de los gonadotrofos y al cese de la secreción de LH y FSH. La célula debe ahora reponer sus receptores y sus reservas de LH y FSH mediante la activación de la síntesis proteica. Esto ocurre en los periodos
entre picos de secreción de LH-RH, durante los cuales los gonadotrofos quedan listos para responder al siguiente pulso de LH-RH.
Los sutiles cambios de secreción de LH y FSH a lo largo de la vida y del ciclo menstrual son promovidos por la unión LH-RH y su receptor, y moduladas por las condiciones intracelulares que afectan a la intensidad de respuesta de las rutas señalizadoras posteriores, cuyos detalles comienzan a conocerse (Fig. 3).

El receptor de LH-RH responde a la estructura típica ce los receptores de siete dominios
hidrofóbicos trasmemebrana, que interactúan con proteínas G. La figura muestra los siete
dominios en dos formas, y la manera en que el péptido de LH-RH establece su unión con el
receptor (parte inferior) contactando con varios de sus dominios.
La infusión continuada de LH-RH desde un dispositivo farmacodinámico apropiado produce un aumento inicial de secreción de LH y FSH, pero en unos días se produce el efecto opuesto, con cese completo de la secreción de gonadotrofinas. Ese estado revierte en cuanto se suprime el aporte continuo de LH-RH, por lo que se le denomina hipofisectomía médica selectiva. La infusión continua de LH-RH produce el cese de la secreción de LH y FSH, lo que da lugar a la ausencia de estimulación ovárica (o testicular: también se usa ese tratamiento en cáncer de próstata), entrando el paciente en la llamada castración química o médica.
La diferencia hormonal entre la ovariectomía (u orquidectomía) y la castración química estriba en que, en el primer caso, la secreción de LH y FSH está irreversiblemente aumentada, debido a la supresión de la realimentación negativa de las hormonas gonadales sobre el eje hipotalámico-hipofisario, mientras que en el segundo caso la secreción de LH y FSH es muy baja, pero reversible tras la supresión de la LH-RH.