2. Ciclo Celular Y Cáncer
2. Ciclo Celular Y Cáncer
Las células normales de cualquier tejido necesitan de estímulos mitogénicos para pasar de un estado quiescente al estado de proliferación activo. Por el contrario, las células tumorales presentan menor dependencia de estímulos externos y se comportan como si se encontrasen continuamente sujetas a un mecanismo de estimulación autocrino. Esta independencia respecto de estimulación externa claramente rompe con las señales que normalmente operan para mantener la homeostasis tisular.
Las estrategias moleculares que permiten a las células tumorales adquirir la autonomía necesaria para su proliferación en ausencia de estimulación externa provienen de señales inapropiadas activadas en la ruta hacia el núcleo a partir de una alteración en: a) la señalización desde la membrana, b) un mediador de la vía a nivel citoplasmático, o c) factores nucleares anormalmente activos (figura 1).
La figura 2 ilustra de forma integrada los diferentes circuitos mostrando las interconexiones que relacionan las vías de señalización celular para los estímulos proliferativos, con las señales que mantienen a la célula en un estado de quiescencia permitiendo su supervivencia, con señales que permiten la comunicación con las células de su ambiente tisular, y con vínculos hacia rutas responsables de inducir la diferenciación celular. Como puede observarse, el resultado del conjunto de señales que convergen sobre una misma célula deben ser canalizadas finalmente hacia la maquinaria de control del ciclo celular.
Como ya se mencionó anteriormente, la maquinaria de control del ciclo celular esencialmente
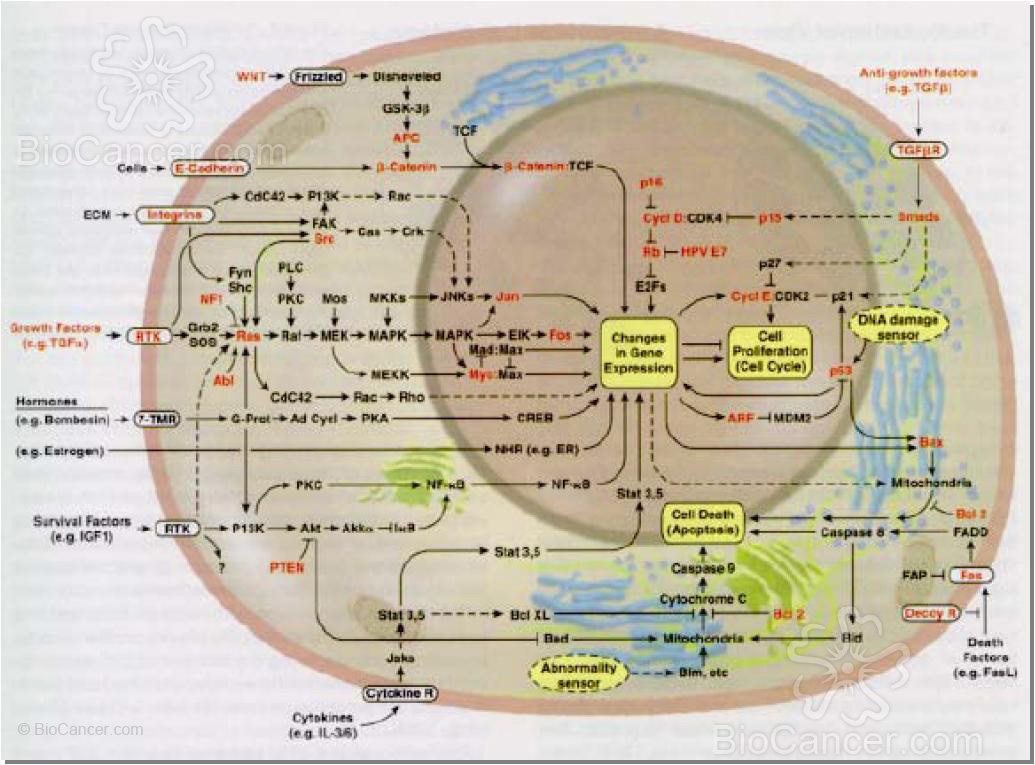
que relacionan las vías de señalización celular para los estímulos proliferativos, con las señales que
mantienen a la célula en un estado de quiescencia permitiendo su supervivencia, con señales que permiten
la comunicación con las células de su ambiente tisular, y con vínculos hacia rutas responsables de inducir
la diferenciación celular. Como puede observarse, el resultado del conjunto de señales que convergen sobre
una misma célula deben ser canalizadas finalmente hacia la maquinaria de control del ciclo celular
está formada por dos tipos de componentes: las cdk's y las ciclinas. El ensamblaje de estas moléculas constituye el ritmo que lleva a la célula a avanzar por las distintas fases de su ciclo (figura 3). Cuando una célula abandona su estado de quiescencia en respuesta a los estímulos mitogénicos, tiene lugar una inducción de la síntesis de ciclinas del tipo D. La continua presencia de señales mitogénicas garantiza que los niveles de estas ciclinas se mantengan suficientemente altos durante el resto del ciclo celular. Una vez sintetizadas estas ciclinas, se produce la formación de complejos con cdk4 y cdk6, dos alternativas cuyas funciones se encuentran por aclarar. En estadios avanzados de la fase G1 se produce la estimulación de la síntesis de ciclina E, que formará complejos ciclina E-cdk2. La activación de estos complejos es clave para la conclusión de esta fase y la entrada en la fase S. Es ya en esta fase cuando se debe expresar la ciclina A, siendo necesaria para que la maquinaria de replicación lleve a cabo la síntesis de ADN. Posteriormente, en la fase G2 se produce una nueva asociación de esta ciclina con otra enzima para formar complejos ciclina A-cdc2, necesarios para iniciar los preparativos de cara a la inminente mitosis.
En esta fase la actividad de la cdc2 es orientada hacia otros sustratos al asociarse con otras ciclinas mediante la formación de complejos activos ciclina B-cdc2.
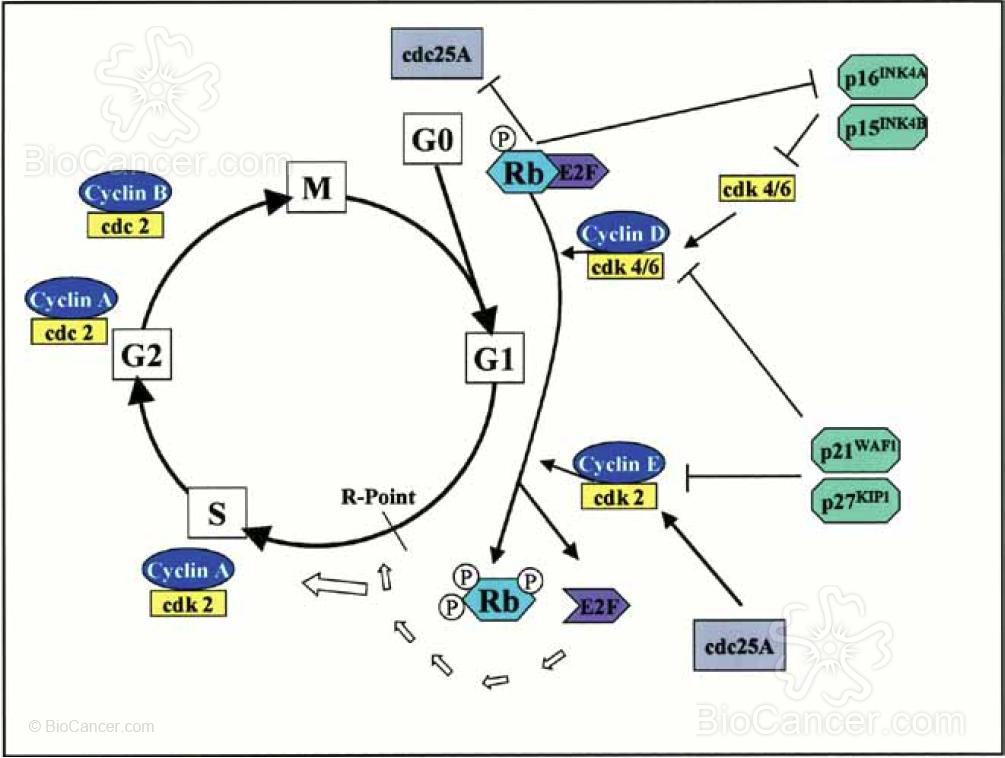
componentes: las cdk’s y las ciclinas. El ensamblaje de estas moléculas constituye el ritmo que lleva a la
célula a avanzar por las distintas fases de su ciclo. Cuando una célula abandona su estado de quiescencia
en respuesta a los estímulos mitogénicos, tiene lugar una inducción de la síntesis de ciclinas del tipo D
que formarán complejos con cdk4 y cdk6. En la fase G1 tardía se produce la síntesis y formación de complejos
c ic lina E-cdk2 cuya activación es clave para la conclusión de esta fase y la entrada en la fase S. En esta
fase es cuando s e expres a ciclina A, siendo necesaria para que la maquinaria de replicación lleve a cabo
la síntesis de ADN. En la fase G2 se producen complejos ciclina A-cdc2, necesarios para iniciar los preparativos
para la inminente mitosis. En esta fase la actividad cdc2 es orientada hacia otros sustratos al formar
complejos activos con otras ciclinas (ciclina B-cdc2).